Histologia: El grup de recerca de Ratul Chowdhury i Rahul Kumar Nelli del College of Veterinary Medicine de la Iowa State University treballa, entre d’altres coses, en virus que afecten la cabana bovina. Ning-Chieh Twu, del mateix grup, és interessat en les interaccions virus-hoste. Com a model en les seves investigacions empren el virus de la grip A, tenint en ment els brots recents de grip aviària H5N1 d’alta patogenicitat que hi ha hagut en granges bovines dels Estats Units. També treballen amb el coronavirus SARS-CoV-2, també reportat en granges bovines. Twu et al. publiquen aquesta setmana a Scientific Reports un estudi que investiga la distribució espacial de receptors d’aquests virus en teixits bovins emprant PixF. PixF és una eina d’anàlisi d’imatges. L’apliquen sobre teixits respiratoris (tràquea i pulmó) i digestius (intestí prim i intestí gros) de bous creuats Holstein-Angus. Amb una tinció histoquímica de lectina amb receptors d’àcid siàlic α2,6-Gal (del saüc Sambucus nigra) i α2,3-Gal (de Maackia amurensis) identifiquen receptors del virus de la grip. Amb immunofluorescència indirecta detecten ACE2. TMPRSS2, APN, DPP4 i CEACAM1, que actuen com a receptors de coronavirus. PixF ofereix un marc de processament d’imatge per a quantificar i mapar l’expressió de receptors. Així, Twu et al. troben un predomini de receptors SA α2,3-Gal en regions epitelials, mentre que els receptors SA α2,6-Gal es troben limitats a teixits glandulars del tracte respiratori. TMPRSS2, APN i DPP4 tenen una alta expressió en l’epiteli de mucoses respiratòries. ACE2, TMPRSS2 i DPP4 tenen una alta expressió en l’epiteli de la mucosa intestinal. CEACAM1 té una baixa expressió general. El més rellevant és que PixF ofereix una informació quantitativa i localitzada de la distribució de receptors virals, essencial per a entendre les interaccions hoste-patogen.
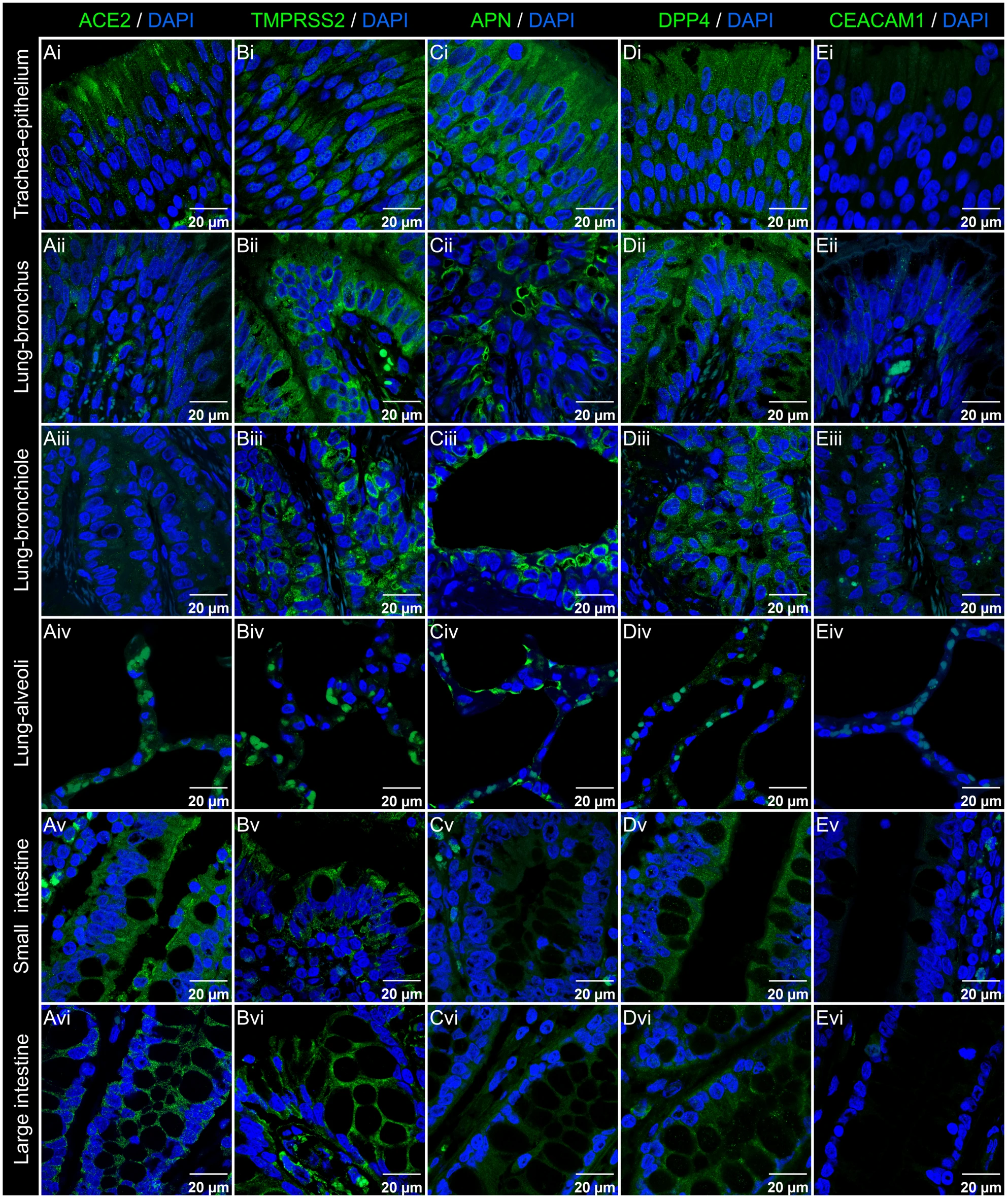
Microfotografies confocals que mostren la distribució de receptors de coronavirus en els tractes respiratori i intestinal de bestiar boví
Receptors virals
Les pandèmies de grip i de malalties coronavirals s’expliquen per l’elevat contagi respiratori en humans d’aquestes infeccions. Molts coronavirus també poden donar lloc a infeccions entèriques, la qual cosa ha estat la base d’epizooties en animals de granja. El concepte d’Una Sola Salut ens recorda permanentment que aquests són virus zoonòtics, que poden mutar i evolucionar ràpidament. Troben reservoris no tan sols en la població humana i en la cabana domèstica, sinó també en la fauna salvatge, terrestre i marina, en mamífers i aus.
Es tracta de virus d’ARN, amb una alta taxa de mutació que s’explica pel procés de replicació genòmica. Un aspecte central en el cicle víric és la interacció entre la partícula vírica i la membrana de cèl·lules susceptibles d’infecció. Els virus de la grip empren com a receptors els àcids siàlics (SA) tant en l’adhesió inicial a la cèl·lula com en l’entrada ulterior.
Els virus de la grip A d’origen aviari s’uneixen preferentment a receptors d’àcid siàlic enllaçats a galactosa per un enllaç α2,3 (SA α2,3-Gal). En pollastres, concretament, la preferència és per a SA α2,3-Gal-β (1–4) N-acetilglucosamina (GlcNAc). En ànecs, en canvi, la preferència és per a SA α2,3-Gal-β (1–3) N-acetilgalactosamina (GalNAc).
Els virus de la grip A d’origen humà o porcí s’uneixen preferentment a receptors SA amb un enllaç α2,6-galactosa (SA α2,6-Gal).
Val a dir que els àcids siàlics també funcionen com a receptors o co-receptors potencials d’altres famílies de virus, com els Coronaviridae, els Paramyxoviridae, els Flaviviridae, els Picornaviridae, els Reoviridae, els Parvoviridae, els Adenoviridae, els Papillomaviridae, els Polyomaviridae, els Caliciviridae, etc.
Els coronavirus, a banda de l’àcid siàlic, empren diversos receptors cel·lulars, com l’ACE2 (enzim convertasa 2 de l’angiotensina), APN (aminopeptidasa N), CEACAM1 o DPP4 (dipeptidil-peptidasa 4). La interacció amb aquests receptors és cabdal per determinar el rang potencial d’espècies on pot circular cada coronavirus. Per això tenen gran interès les tècniques de tinció i imatge que posen de manifest la distribució d’aquests receptors virals en cèl·lules i teixits de l’hoste. La tinció dual amb tints fluorescents pot ajudar a fer anàlisis de co-expressió. La microscòpia confocal forneix resolucions d’imatge per a aquests senyals fluorescents (habitualment d’emissió verda i vermella). L’anàlisi píxel a píxel pot identificar els casos de veritable co-localització dels dos senyals.
Twu et al. han desenvolupat una eina d’anàlisi de fluorescència basada en píxels per a una ràpida caracterització d’imatges. L’anomenen ‘PixF’, i amb ella mesuren la intensitat espacial de l’emissió vermella i verda de cada punt de la imatge. PixF ofereix una visualització en tres dimensions, on els eixos x i y representen el pla del tall histològic i z és el valor d’intensitat absoluta de la fluorescència.
Twu et al. són conscients que PixF no és capaç de suplir tota la funcionalitat de plataformes com ImageJ, CellProfiler o Imaris. L’objectiu de PixF és la velocitat i l’accessibilitat que pot oferir en segons sobre centenars d’imatges.
La plataforma PixF
Twu et al. recolliren mostres de tràquea, pulmó, intestí prim i intestí gros de tres bous de tres anys del creuament Holstein-Angus sacrificats en el Iowa State University Meat Laboratory. Els teixits eren fixats en 10% de formalina neutra tamponada i inclosos en blocs de parafina. Els talls de 4 μm eren transferits a portaobjectes i preparats per a anàlisi.
La tinció amb lectina posa de manifest els àcids siàlics:
- la lectina SNA de Sambucus nigra interactua amb α2,6 SA.
- la lectina MAL-I de Maackia amurensis interactua amb SA α2,3-Gal-β (1–4) GlcNAc.
- la lectina MAL-II de Maackia amurensis interactua amb SA α2,3-Gal-β (1–3) GalNAc.
Prèviament els portaobjectes eren desparafinats i blocats amb biotina i streptavidina. Les lectines eren marcades amb isotiocianat de fluoresceïna (FITC).
L’assaig d’immunofluorescència per a la detecció de SA es feia amb un anticòs policlonal anti-Neu5Gc.
L’ús d’anticossos específics marcats amb fluorocrom (immunofluorescència indirecta) posa de manifest receptors de coronavirus.
Els portaobjectes, després de la immunotinció, eren mantinguts en la foscor durant 24 hores, i llavors se’n prenien imatges amb un microscopi confocal.
En l’anàlisi PixF hi havia un pas inicial de definició dels mapes de color que tradueixen la fluorescència a l’escala RGB. Els canals vermell i verd reporten els senyals dels dos fluorocroms emprats, i el canal groc reporta la superposició dels dos senyals.
La distribució de l’àcid siàlic en el tracte respiratori boví
En l’epiteli del tracte respiratori trobem un predomini de receptors SA α2,3-Galβ (1–3). Això el fa potencialment susceptible a virus de la grip aviària. Troben aquests receptors especialment en la membrana apical de cèl·lules de l’epiteli pseudoestratificat ciliat de tràquea i bronquis, però també en bronquiols i alvèols.
Els receptors SA α2,6-Gal es troben relativament confinats en les cèl·lules calciformes (secretores de mucus), en les cèl·lules basals i en les glàndules submucoses. Aquests són els receptors típics de virus de la grip A d’origen mamífer.
La distribució d’àcid siàlic en el tracte intestinal boví
En el tracte intestinal boví hi ha una expressió abundant de receptors SA α2,3-Gal. En canvi, SA α2,6-Gal es troba relativament restringit a la regió subepitelial i a les cèl·lules calciformes.
La distribució de receptors de coronavirus en els tractes respiratori i intestinal
Hi ha una forta expressió d’ACE2 en l’epiteli de la tràquea, dels alvèols i dels budells. És menor aquesta expressió en l’epiteli de bronquis i bronquiols.
TMPRSS2, APN i DPP4 són àmpliament expressats en l’epiteli respiratori, i especialment en els alvèols. En l’epiteli intestinal, TMPRSS2 i DPP4 tenen una expressió uniforme, tant en la regió de les vellositats com en les criptes. CEACAM1 és present en el tracte respiratori però no se’l detecta en el tracte intestinal.
L’aplicació de PixF en predicció virològica
El bestiar boví és susceptible als virus de la grip C i grip D, al coronavirus boví (BCoV) i al SARS-CoV-2. Aquests virus poden produir afectacions respiratòries i gastrointestinals.
L’àcid siàlic és el receptor que permet l’adhesió i entrada del virus de la grip A. Però també són molts altres els virus, bacteris, fongs i paràsits que hi interactuen.
Entre el virus de la grip A es distingeixen habitualment entre els virus d’origen aviar i equí, que tenen una afinitat preferent per SA α2,3-Gal, i els virus d’origen humà i porcí, que tenen una afinitat preferent per SA α2,6-Gal. Els porcs, amb presència simultània dels dos receptors, poden actuar com a reservori de contacte entre els dos tipus de virus de grip A. Ara bé, Twu et al. ens recorden que també en teixits bovins coexisteixen els dos tipus de receptors, la qual cosa fa que també la cabana bovina sigui un lloc potencial de recombinació de virus de grip A d’origen diferent.
Cal recordar, però, que la distribució de receptors cel·lulars varia segons l’edat, sexe, raça i estat de salut de cada animal, i que és alhora específica de teixit. A això hem d’afegir l’efecte del tipus d’anticòs, temps d’incubació i tècnica d’imatge per visualitzar-los.
PixF es troba actualment en l’estadi de versió beta. Té aplicacions potencialment en la patologia i radiologia diagnòstiques. El Laboratori de Chowdhury la posa en forma de pàgina web a disposició d’investigadors i clínics. Confien que també pot ésser una eina d’aprenentatge per als qui s’inicien en l’àmbit de processament de microfotografies d’immunofluorescència.
Lligams:
- Spatial mapping of influenza and coronavirus receptors in the respiratory and intestinal tract epithelium of beef cattle using advanced PixF image analysis. Ning-Chieh Twu, Yee Chuen Teoh, Curwen Pei Hong Tan, Riza Danudoro, Xiaoyi Cheng, Raquel Espin-Palazon, Todd Bell, Luis Giménez-Lirola, Ratul Chowdhury, Rahul Kumar Nelli. Scientific Reports 15: 44029 (2025).
- Pixel Analytics Suite (PixF), de Chowdhurylab.